
| |
| Keap1-Nrf2制御系の活性制御メカニズム |
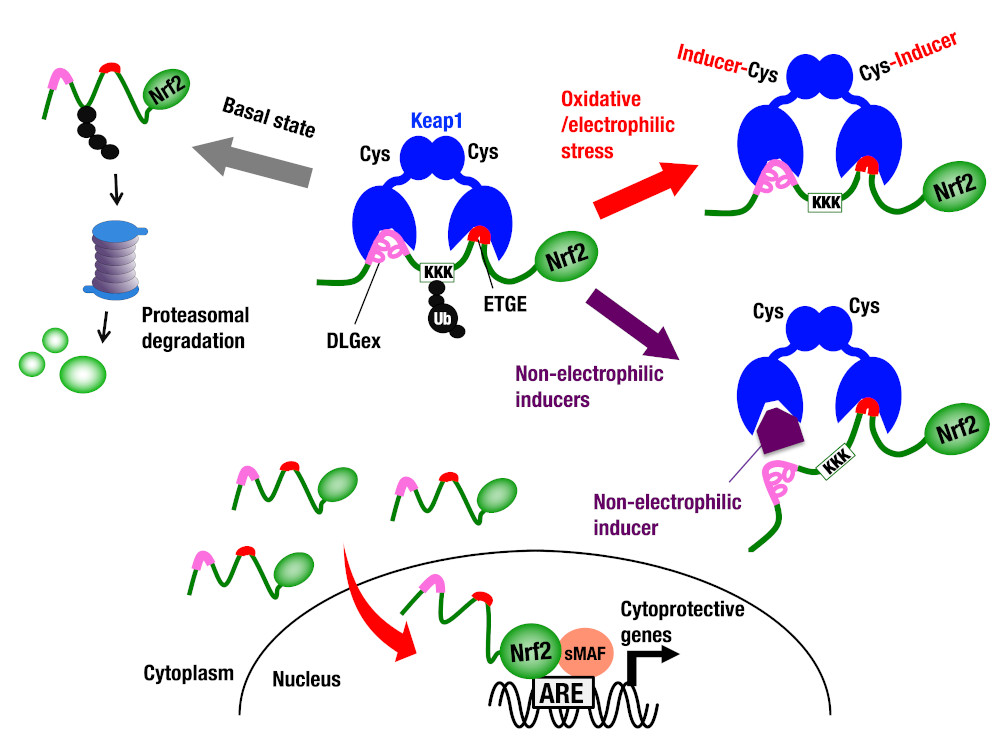
| 細胞が酸化ストレス・親電子性物質にさらされると,グルタチオン合成酵素やヘムオキシゲナーゼ1などの抗酸化蛋白質や異物代謝酵素を発現誘導することで生体防御に努めます。この酸化ストレスによる遺伝子発現機構に関しては,遺伝子上流に存在する抗酸化剤応答配列ARE(antioxidant response element)を介した転写レベルの発現調節が重要です。この制御配列には,塩基性ロイシンジッパー(bZip)型転写因子であるNrf2と小Maf群因子によるヘテロ二量体が結合し,遺伝子発現を強力に活性化します。非酸化ストレス下では,Nrf2はKeap1と名づけられた因子により細胞質に留められ、プロテアソーム依存的なタンパク質分解抑制を受けています。細胞に酸化ストレスないし親電子性物質がさらされると,Keap1によるNrf2抑制機構は解除され,核移行したNrf2が防御遺伝子の発現を誘導します。すなわち、Keap1-Nrf2システムは,ストレス応答型の転写制御システムを形成しているのです。 |
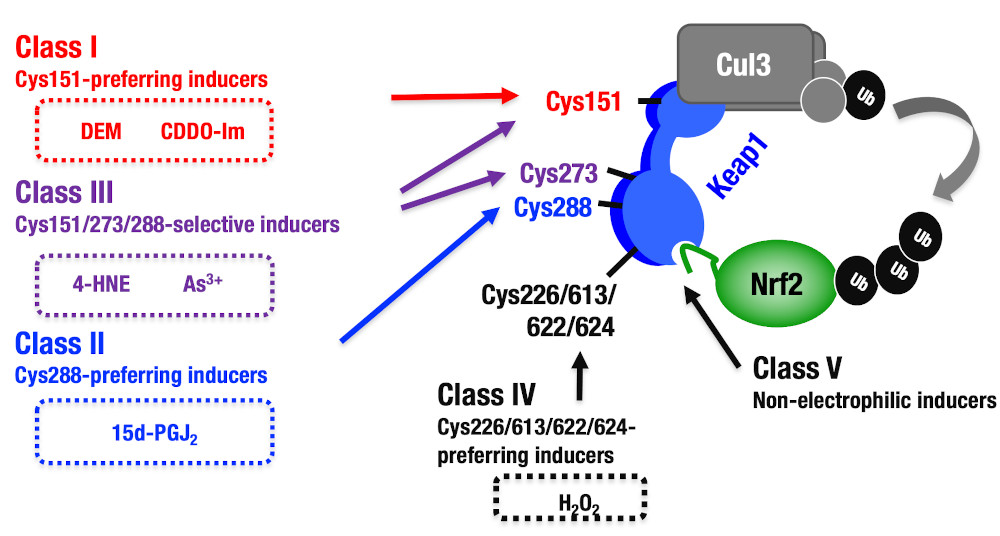
| Keap1は複数のシステイン残基を使い分けることにより酸化/親電子性ストレスを感知しています。本研究を進めていくことで、Keap1−Nrf2システムの活性化メカニズムの分子基盤の理解が進み、Keap1−Nrf2システムを利用した新薬や治療法の開発が加速することが期待されます。 Suzuki et al, 2019 Cell Reports |
| 参考文献 Suzuki T, Hidaka T, Kumagai Y, Yamamoto M. (2020) Environmental pollutants and the immune response in press. Nature immunology. 21, 1486-1495. Suzuki T, Muramatsu A, Saito R, Iso T, Shibata T, Kuwata K, Kawaguchi S, Iwawaki T, Adachi S, Suda H, Morita M, Uchida K, Baird L, Yamamoto M. (2019) Molecular mechanism of cellular oxidative stress sensing by Keap1. Cell Reports 28, 746-758. Suzuki T, Seki S, Hiramoto K, Naganuma E, Kobayashi HE, Yamaoka A, Baird L, Takahashi N, Sato H and Yamamoto M. (2017) Hyperactivation of Nrf2 in early tubular development induces nephrogenic diabetes insipidus. Nature Communications. 8, 14577. Suzuki T, Yamamoto M. (2017) Stress-sensing mechanisms and the physiological roles of the Keap1-Nrf2 system during cellular stress. Journal of Biological Chemistry 292, 16817-16824. Kobayashi HE, Suzuki T, Funayama R, Nagashima T, Hayashi M, Sekine H, Tanaka N, Moriguchi T, Motohashi H, Nakayama K, and Yamamoto M. (2016) Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nature Communications. 7, 11624. Saito R, Suzuki T, Hiramoto K, Asami S, Naganuma E, Suda H, Iso T, Yamamoto H, Morita M, Baird L, Furusawa Y, Negishi T, Ichinose M, Yamamoto M. (2016) Characterizations of Three Major Cysteine Sensors of Keap1 in Stress Response. Molecular and Cellular Biology. 36, 271-284. Iso T, Suzuki T, Baird L, Yamamoto M. (2016) Absolute amounts and status of Nrf2-Keap1-Cul3 complex within cells. Molecular and Cellular Biology, 36, 3100-3112. Suzuki T, Yamamoto M. (2015) Molecular basis of the Keap1-Nrf2. Free Radical Biology and Medicine. 88, 93-100. Suzuki T, Shibata T, Takaya K, Shiraishi K, Kohno T, Hunitoh H, Tsuta K, Furuta K, Goto K, Hosoda F, Sakamoto H, Motohashi H, Yamamoto M. (2013) Regulatory nexus of synthesis and degradation deciphers cellular Nrf2 expression levels. Molecular and Cellular Biology. 33, 2402-2412. Suzuki T, Motohashi H, Yamamoto M. (2013) Toward clinical application of the Keap1-Nrf2 pathway. Trends in Pharmacological Science 34, 340-346. Yamamoto T, Suzuki T, Kobayashi A, Wakabayashi J, Maher J, Motohashi H and Yamamoto M. (2008) Physiological Significance of Reactive Cysteine Residues of Keap1 in Determining Nrf2 Activity. Molecular and Cellular Biology. 28, 2758-2770. Suzuki T, Kelly VP, Motohashi H, Nakajima O, Takahashi S, Nishimura S and Yamamoto M. (2008) Deletion of selenocysteine tRNA gene in macrophage and liver results in compensatory gene induction of cytoprotective enzymes by Nrf2. Journal of Biological Chemistry. 283, 2021-2030. 私たちのグループでは、Keap1-Nrf2システムによるストレス応答メカニズム解明を目指しています。 私たちと一緒に真剣にサイエンスに取り組んでみたい方は、 鈴木隆史(taka23@med.tohoku.ac.jp)までお問い合わせください。 |
| プロジェクト一覧へ戻る | [編集] |